


1ste manier: vertaling etc volgt nog (ben zelf nog effe info aan het verzamelen enzo, die smijt ik eerst hierop en dan verwerk ik da wel verder tot een deftig topicje ") )
)
Vrouwelijke zaden kweken
Gefeminiseerde / vrouwelijke zaden creëren is een kunst, er zijn enkele verschillende manieren. Je hebt glibberic Acid, licht stress, ph-stress, stress door je meststoffen maar allemaal willen ze een ding bereiken dat er mannelijke bloemen op een XX plant komen.
Dit proces kan toegepast worden om:
A: nieuwe vrouwelijke zaden te creëren van een supermoeder die je staan hebt
B: om interessante vrouwelijke hybride zaden te creëren van verschillende interessante soortjes die je momenteel hebt
C: om vrouwelijke zaden te creëren voor een optimale buitenkweek
D: om het proces te versnellen naar de zoektocht voor een ideale nieuwe moederplant
E: om het aantal planten die je kweekt voor medicinaal gebruik zo beperkt mogelijk te houden (dus geen extra planten uit angst voor teveel mannetjes)
F: voor meer variatie, door stabiele vrouwelijke zaadlingen te maken die kunnen dienen als een alternatief voor klonen
Het is wel een vereiste dat je je al goed ingestudeerd hebt over de theorie van cannabis kweken voor dat je een techniek als deze gebruikt.
Het is ook zeer belangrijk dat je de basis veiligheidsvoorschriften respecteert wanneer je de chemicaliën mixt en gebruikt. De risico`s zijn gelijk aan het mixen en gebruik van chemische meststoffen dus pas dezelfde veiligheids voorzorgen toe.
Onthoud: niets zal nooit goeie genetica vervangen, en een deel van wat je kweekt zou altijd moeten terugkeren op de professionele cannabis kweker... Diegene die vele generaties gewerkt heeft om van een soortje uiteindelijk hun echte F1 meesterwerkjes te bekomen. Help hen door hun zaden te kopen zo kunnen zij verder werken aan nieuwe sterke soorten voor ons thuiskwekers..
-------------------------------------
1 STS
Preparation of STS:
Eerst word een voorraad oplossing gemaakt. Het bestaat uit 2 delen (A en B) die eerst apart worden vermengd en uiteindelijk dan samen vermengd.
Deel A wordt altijd samen gemixed met deel B terwijl je snel roert. Gebruik gedestilleerd water, kraantjeswater kan precipitaten vormen.
Draag handschoen tijden het mixen en gebruiken van deze chemicaliën, en mix en gebruik deze enkel in een deftig geventileerde ruimte. Een masker kan voorkomen dat je een bepaalde stof inademt, die agressief is. STS is kleurloos en geurloos, en betekent een minimum aan risico als je alles toepast zoals hier boven beschreven. Onthoud dat silvernitraat en sts bruine plekken kunnen veroorzaken bij het indrogen, dus spuit over krantenpapier en vermijd morsen.
Deel A: 0.5 gram zilver nitraat vermengd met 500ml gedestilleerd water
Deel B: 2.5 gram natriumthiosulfaat (watervrij) vermengd met 500ml gedestilleerd water
Het zilvernitraat lost op binnen de 15 seconden. Het natriumthiosulfaat heeft ongeveer 30/45 seconden nodig om op te lossen.
De zilvernitraat oplossing (deel A) word dan vermengd met het natriumthiosulfaat oplossing (deel B) terwijl je goed en snel roert. Het resultaat is een voorraad zilver thiosulfaat oplossing (STS).
Deze voorraad oplossing wordt vervolgens verdund in een verhouding van 1:9 om een werkende oplossing te maken. Bijvoorbeeld: 100ml STS is toegevoegd aan 900ml gedestilleerd water. Deze oplossing wordt dan verneveld op de geselecteerde vrouwelijke planten.
De beide producten de voorraad STS zowel als de gemaakte oplossing word best in de koelkast bewaart na gebruik, om activiteits verlies te voorkomen. Teveel aan oplossing kan altijd veilig door het wc gespoeld worden of enige andere afvoer met stromend water met verwaarloosbare gevolgen voor het milieu.
-------------------------------------
Gebruikswijze:
De STS oplossing wordt dan besproeid op de vrouwelijke planten tot het er mooi afdruipt. Doe het sproeien over een krant in een aparte ruimte. Je zal waarschijnlijk niets ruiken maar ventileer toch maar. Nu heb je wat ik een "F>M plant" noem; een vrouwelijke plant die mannelijke bloemen gaat produceren.
Nadat de F>M plant droog is, plaats je ze direct onder 12/12. Dit wordt meestal 3/4 weken voor de doel datum (om te bestuiven) wanneer de planten klaar zijn om bestuift te worden. Responstijden kunnen enigszins variëren, afhankelijk van de soort. Meer specifieke tijden kunnen worden bepaald door met uw eigen strain proeven te doen. In mijn onderzoek duurde het 26 dagen voor de eerste pollen. 30-35 dagen lijkt een optimale planning.
Zo, veronderstel dat je doelplant 3/4 weken nodig heeft om volledig volwassen zaden te produceren, een strain dat 8 weken nodig heeft om volwassen te worden moet in bloei gezet worden op ongeveer dezelfde tijd als de F>M plant. Een doelplant dat afbloeit in 6weken moet dan 10 dagen later in bloei gezet worden zodat ze niet al afgebloeid is voor de zaden volledig volwassen zijn.
Een afzonderlijke tak die je bestoven hebt mag je altijd iets langer volwassen laten worden aan een plant, je moet ze niet meeoogsten samen met de zaadloze toppen. Laat enkel genoeg bladeren aan de plant zodat ze gezond kan blijven.
-------------------------------------
Effecten:
In enkele dagen zag ik al bladeren vergelen aan de F>M planten. Dit verschijnsel bleef ongeveer 2 weken duren: daarna werden ze langzaam terug groen, buiten enkele grote fan bladeren. Voor de rest leek de plant gezond. Geen verbranding te zien. De groei stopte wel voor 10 dagen, en starte erna terug langzaam. Geen strekken te zien. Na 2 weken vormden de F>M planten duidelijke mannelijke bloemen. Niet enkel een paar ballen maar echte complete mannelijke toppen. 1 plant vormde nog vrouwelijke bloemen, maar in het geheel bekeken was deze overwegend mannelijk.
Het is vreemd om een oude vriendin die je als de achterkant van je hand kent te zien een seksverandering te ondergaan.
Als de F>M planten eruit zien alsof zij snel kunnen openen pollen laten vrijkomen, (3-1/2 tot 4 weken) verplaats je hen best naar een aparte ruimte (weg van je bloeiruimte) met licht op 12/12. Je moet niet teveel inzitten over wattages; het is maar tijdelijk.
Als de pollen vrijkomen verplaats je best je doelplanten bij in de kast zodat ze deze kunnen bestuiven.
Een meer gecontroleerde aanpak is je F>M planten te isoleren in een derde afzonderlijke ruimte(geen licht is hier nodig, deze zijn nu toch pollen aan het lossen dus zijn ze toch op hun einde). In deze andere ruimte kunnen we de pollen voorzichtig verzamelen met een plastiekzak of een vel krantenpapier. Dan kunnen we onze doelplanten in de bloeiruimte op deze manier bevruchten. Als je het zo doet let wel op dat je pollen van je F>M plant niet mixen of dat ze niet kruis bestuiven. Dus vermijd beweging.
Let er speciaal op dat deze pollen zich niet verzamelen buiten aan de buitenkant van je zak door statische elektriciteit bv.
Neem kleine open clusters van de binnen kant van de bloemen in de zak en schud ermee. Ga dan best naar buiten om de overtollige lucht uit de zak vrij te laten. Let hierbij op dat er geen pollen op je kleren of handen terecht komen.
Nu kan je deze zak voorzichtig over een tak schuiven en dan afbinden rond de tak met een stukje tape of koord, zodat de pollen bij de top opgesloten zit. Laat de zak zich langzaam opvullen met een beetje lucht voor je hem afsluit aan de tak, zodat de tak nog kan "ademen". Deze techniek zorgt ervoor dat je bij een plant verschillende takken kan bestuiven met verschillende pollen van verschillende strains of dat je een deel van de plant ook nog kan oogsten als sensemilla.
Na 4-5 dagen mag je de zak voorzichtig verwijderen en de plant kan zen bloeicyclus verder vervolledigen.
Maar gelijk welk van deze wijzes dat je toepast uiteindelijk heb je na 2/4weken rijpen vrouwelijke zaden over.
-------------------------------------
Over de chemicalieen:
Zilvernitraat is een witte kristallijne lichtgevoelige chemische stof die gewoonlijk wordt gebruikt in de fotografie. Het wordt ook gebruikt in de ogen van baby`s bij de geboorte om blindheid te voorkomen. Het kan leiden tot lichte irritatie van de huid, en bruine vlekken. Vermijd inademen. Ik heb geen geur of rook ondervonden maar ventilatie word aanbevolen. Een halve gram STS kost verrassend weinig en zou passen in een gel-capsule.
Have fun met het testen van deze techniek. Gebruik het met verstand. En bovenal gebruik STS bij kwaliteits F1 zaden geproduceerd door een professionele kweker.
2 Rodelization (soma)
In mijn zoektocht naar mooiere milieuvriendelijke manieren voor te werken met cannabisplanten, heb ik een nieuwe manier ontdekt om vrouwelijke cannabiszaden te ontwikkelen. Vrouwelijke zaden ontstaan door stressen niet door genetica. Alle cannabisplanten kunnen en zullen mannelijke bloemen produceren bij stress. Sommige soorten hebben liever een hogere P, sommige een lagere. Sommigen hebben veel voeding nodig anderen dan weer minder. Daar zit veel variëteit in de cannabis genetica, en je kan niet elke plant gelijk behandelen.
Als je een bepaalde soort wilt leren kennen neemt het echt een groot aantal kweken in beslag voor je je echt kan zeggen dat je die soort kent. net als je een vriend wil leren kennen, dit neemt veel tijd in.
Ik groei nu al verschillende strains voor meer dan een decennium en ik begin echt alle nuances te kennen die verschillende planten bezitten. Ik kan ze herkennen van op een afstand. Ik moet wel zeggen dat ik hierbij veel hulp krijg van mijn vrienden, zowel voor het maken van zaden als voor het beter leren kennen van deze heilige plant.
Ik noemde deze nieuwe methode 'Rodelization' maar een vriend die me hielp te realiseren en gebruik te maken van deze manier om vrouwelijke zaden te maken. Nadat ik dezelfde soort kweekte oogst na oogst onder dezelfde condities, merkte ik dat als ik deze planten 10 tot 14 dagen langer liet staan dan gewoonlijk dat deze mannelijke "bananen" vormde. Een mannelijke banaan is een zeer kleine mannelijke bloem die zich vormt door stress. Normaal laten zij geen pollen vroeg genoeg los om cannabis zaden te maken, maar soms wel. Dit is een soort ingebouwde veiligheid voor de plant, zodat de plant zeker is dat zij zich kan voortplanten ook in extreme condities.
Voor mij is een mannelijk bloem een zeer mooi gebeuren. Het geeft de mogelijkheid om vrouwelijk zaad te maken. Veel kwekers hebben een natuurlijke angst voor mannelijke bloemen 'bananen'. Ze zien een bloem en willen al direct hun hele oogst weggooien of gaan er met een pincet angstwekkend rond de planten om deze banaantjes te plukken. Ik noem deze 'veiligheids voorzieningen' omdat deze voorkomen bij stress.
Gebruikswijze:
Bij de Rodelizaion methode is de mannelijke banaan zeer waardevol. Nadat je je vrouwelijke plant 10/14 dagen langer dan normaal hebt laten bloeien, hang ze op om te drogen, neem ze nadien voorzichtig van de drooglijnen en inspecteer voor bananen. Elke banaan zou moeten verwijdert worden en voorzichtig in een kleine zak geplaatst worden. Deze afgesloten zakken kan je in de frigo bewaren voor 1 tot 2 maand en ze zullen nog steeds potent zijn.
Voor de tweede fase heb je een oogst nodig dat al 2.5 weken in bloei staat. Neem je gesloten zak uit de frigo, en vervolg je plan om de vrouwelijke toppen te bevruchten. Om dit te doen, moet je eerst de vrouwelijke plant en de pollen van de vorige strain uitkiezen. Dan zet alle vents en fans uit in de ruimte. Neem een kleine verfborstel, dip het in de pollen, en verf het op de vrouwelijke toppen. Doe dit voor iedere strain die je wil samen groeien. Ik heb dit gedaan met 10 verschillende strains in dezelfde ruimte met groot succes.
Ik gebruik voornamelijk de lager gelegen takken om weed zaad van te maken, de topcola`s laat ik zaadloos om te roken. Deze methode neemt wel tijd in (2oogsten), maar is volledig organisch en zorgt ervoor dat je mooie kwaliteit rookbare weed ook hebt en tegelijkertijd maak je ook vrouwelijke zaden aan. Als je een van die kwekers bent die nooit zaden heeft durven kweken uit angst om niets goed over te hebben om te roken, dan zal je zekers houden van deze manier van werken.
Je kan natuurlijk ook deze pollen gebruiken om nieuwe kruisingen vrouwelijke zaden te maken, door kruisbestuiving. De oudere vrouwen met de banaantjes kan je ook in de kamer met de jongere vrouwtjes zetten als ze ongeveer 3 weken in bloei zijn. Zet de circulatie fans vollen bak, en de kleine beetjes pollen vinden hun weg wel terug door de kweekruimte. Houdt dit enkele dagen vol en 6 tot 7 weken later heb je 100% rijpe vrouwelijke zaden; niet zoveel als er zouden gemaakt zijn moest er een mannelijk plant bijgestaan hebben maar genoeg om opnieuw te starten ergens met dezelfde genetica.
Soma
3 Ethyleen gas
Behandelen van cannabis zaad met ethyleen gas zorgt ervoor dat de kans op meer vrouwelijke planten verhoogt met ongeveer 50% (volgens sommigen zelfs 100%, wil ik wel es testen desnoods met hempzaad). Ethyleen word geproduceerd door verschillende planten (bv. bananen, komkommers en meloenen), en deze kunnen op een gemakkelijke manier gebruikt worden om cannabiszaad te behandelen.
Gebruikswijze:
Ongeveer 2 weken voor je van plan bent om je zaadjes te ontkiemen, steek ze in een papierzak of envelop en steek deze in een plastiek zak samen met de bananenschillen van een nog rijpende banaan of komkommer. Vervang de schillen om de paar dagen samen met de papieren zak zodat je schimmel voorkomt.
4 Colloïdaal Zilver
Wat is Colloïdaal Zilver?
Colloïdaal zilver is zuiver, metallic zilver (het element), met deeltjes van 15 atomen of minder, elk met een positieve elektrische lading en bevestigd aan een molecuul van een eenvoudig eiwit. Deze elektrisch geladen deeltjes van zilver zijn uiterst klein, meestal variërend van ongeveer 0.001 tot ongeveer 0,01 micron in diameter, en zijn gesuspendeerd in gedeïoniseerd water. De kracht van de elektrische lading is sterker dan de kracht van de zwaartekracht, dus de zilveren deeltjes blijven opgeschort.
Hoe wordt het toegepast?
De zilverionen in colloidaal zilver reageert als ethyleen remmer/antagonist die de plant forceert om mannelijke bloemen aan te maken die pollen bevatten die enkele vrouwelijke chromosonen bevatten.
Quote:
Treatments which reduce the ethylene level in the tissues (hypo-baric conditions, treatment with benzothiodiazole) or antagonize the action of ethylene (CO2) cause the formation of male or bisexual flowers in place of female ones (Byers et al. 1972)
Wij gebruiken eenvoudigweg een normale lege spuitbus om het op de bladeren te sproeien van een geweten vrouwelijke plant om de pollen op te wekken. Dit kan je doen Met als resultaat 'gefiminiseerde pollen' in tegenstelling tot het regulier stuifmeel bij een man. Als gevolg hiervan, is elk vrouwtje bestoven met de gefeminiseerd pollen een producent van uitsluitend vrouwelijk zaad. We kunnen hier zelfs een moeder voor gebruiken van een 'CLONE-ONLY' strain, zelfs een vrouwelijke plant gegroeid uit feminised zaad. Het enige dat van belang is dat dit een vrouwelijke plant is.
aanrader om te lezen
Induction of Fertile Male Flowers in Female Cannabis (Uni paper on feminised seeds)
H.Y. Mohan Ram and R. Sett. Department of Botany, University of Delhi, Delhi (India)
Beetje uitgebreider:
-Wanneer start ik best met sproeien
Je begint best 2 weken voor de planten op 12/12 gaan en blijven sproeien tot je pollen ziet ontwikkelen.
-Hoe lang sproeien
Normaal gezien sproeien we ongeveer 2 weken.
(eenmaal de plant besproeit, is ze niet langer rookbaar. Dus niet oproken!!)
-Je kan aan zelfbestuiving doen
Dit gebeurt ook in de natuur bv. als een plant zeer oud word (rhodelization), waarbij een vrouwelijke plant enkele bananen afgooit om zichzelf te bestuiven. Dit is een soort van species survival mechanism veronderstel ik.
Je verkrijgt wel niet veel zaad op deze manier en sex tussen twee personen is de norm in het leven, dus persoonlijk probeer ik altijd een 2e vrouwtje als doel voor de mannelijke pollen in de ruimte te plaatsen, en gebruik ik zelfbestoven planten enkel als ik geen andere optie heb.
Ik vermoed dat mss de zelfbestoven zaden kunnen uitgroeien als een kloon-a-like? Maar daar ben ik niet zeker van.
-Op welke mogelijke manieren kan dit allemaal falen
Collodiaal Zilver is zo makkelijk om te maken, gebruiken, en het geeft ons de mogelijkheid om zelfs de nmoeilijkste soortjes te forceren, dus het volledige proces is eigenlijk zeer eenvoudig en zeer effectief als je de basis volgt.. Maar als je dit niet doet, kan het volledig verkeerd zijn.Als je ooit al es CS gebruikt heb maar het niet gelukt is om een plant te reversen, dan is er een goeie kans dat je simpelweg een foutje gemaakt hebt in de procedure.
-De hoofdproblemen
- geen gedestilleerd water gebruiken
- geen zuiver zilver gebruikt 999.999
- de elektroliese niet lang genoeg laten duren
-sproeien met een niet sterk genoege oplossing
-niet genoeg gesproeid (de plant moet volledig doorweekt zijn)
-niet regelmatig genoeg gesproeid
-niet lang genoeg gesproeid (sommige soortjes moet je wel es 3 weken sproeien)
Hoe maak je colloidaal zilver?
Benodigdheden voor de colloidaal zilver generator:

1)een DC adapter (9V/ 300mA-800mA).(600mA is optimaal) gedestilleerd water
2)1/2 quart mayonnaise bokaal 2x 99.99% pure zilver draden , munten, of een zilver bar
3)aquarium luchtpompje
4)2 geisoleerde alligator clips
5)houten plankje van 10cm x 20cm
6)zaag of zwitsers zakmes
7)houtlijm
8)elektrische boormachine en bit
9)een plastiek deksel voor de mayonaise pot of een rubber elastiek ofzo
10)een stukje hout
Maken van de generator

1)Zaag je houten plankje zodanig dat de bokaal en de adapter erop passen
2)Zaag het ander houten latje zodanig dat je het van boven over de mayo pot kan plaatsen. Boor 2 gaten 2 cm van elkaar (om de zilverdraden op te houden). Die gaten zouden iets groter moeten zijn dan de diameter van de zilverdraden. Omdat we daar dan die draden gaan doorsteken. Lijm 2 smalle houten plankjes op de uiteinden van het houten latje, zodat het latje past op de bovenkant van de mayonaise pot
Indien je zilver stukken of bars gebruikt, moet je dit design wel een klein beetje aanpassen.
3)Knip bij de AC adapter de connector af, niet te kort zodat je toch nog beetje draad over hebt. Strip een 2 tal cm draad af
4)Bevestig de gestripte draad aan de aligatorklemmetjes
5)Monteer de generator zoals op de foto
6)Vul de bokaal met gedestilleerd water tot een paar cm van de top
7)Sluit de 2 alligator klemmen aan op de zilver draden die uitsteken uit de houten lat, en steek ze door de geboorde gaten in de houten lat op de bokaal.
8)Sluit de adapter en luchtpomp aan op een timer. Nu ben je klaar om de CS oplossing te maken.
9)Zet de timer op 8 tot 20uren (8u zou genoeg moeten zijn, maak dat je +30PPM hebt in je oplossing
10)Let erop dat de pomp en de ac adapter werkt. De pomp kan je horen maar de DC output ga je moeten testen met een voltmeter. De bubbels zorgen ervoor dat de kwaliteit verbeterd van de zilver oplossing. Kleine fuzzy bubbels zullen zich vormen in 1 pool. Een zaklamp kan je helpen om dit te zien.
11)Nadat dit proces afgelopen is, zeef je je oplossing door een niet gebleekt koffiefilter om kristallen en witte schilfers te verwijderen. Bewaar het elexir in een donkere glazen fles op een koele plaats. Je kan deze enkele maanden bewaren maar het is wel best dat je kleine hoeveelheden maakt zodat je het steeds vers kan gebruiken.
12)Was de mayonaise bokaal en luchtpomp na ieder gebruik en dan spoelen met gedestilleerd water. Reinig de zilverdraden met een scotchbrite pad tegen onzuiverheden en zilverkristallen, spoelen erna.
13)Bewaar nadien alles samen op een stofvrije plaats.

nog enkele manieren om een generator te maken + gebruikswijze: http://www.quantumbalancing.com/makeyourowncs.htm
Waarom CS gebruiken?
Goed, u kunt ook gebruik maken van zilver theosulphate (= nitraat + natrium theosulphate), zilvernitraat (zilver + salpeterzuur), of gibberellinezuur, die allemaal succesvol zijn gebleken in deze taak, maar CS is niet giftig , niet-bijtende, gemakkelijk en veilig en goedkoop te maken thuis, vereist geen DEA aanvraagformulier, en is niet een gereguleerde stof. U kunt het zelfs kopen bij uw plaatselijke drogist / apotheek (sommige mensen drinken het voor haar veronderstelde voordelen voor de gezondheid), en het is goedkoop.
Voor mij, is colloïdaal zilver ongeëvenaard. Colloïdaal zilver werkt als ethyleen remmer / antagonist, wat ons toelaat om het proces te forceren op elk moment we kiezen, zelfs op hard stress-resistente planten (CS maakt geen gebruik van stress om zen magie uit te voeren).
5 Gibberellinezuur
Wat is Gibberellinezuur?
Gibberellinezuur (GA3) is een plantenhormoon, dat de groei en strekking van plantencellen bevordert. Ook beïnvloedt het de verhouding tussen mannelijke en vrouwelijke bloemen op een plant, bevordert het de vruchtzetting, geeft het grotere bladeren en kan sommige tweejarige planten al in het eerste jaar laten bloeien. Verder stimuleert het de cellen van kiemende zaden om mRNA coderend voor hydrolytische enzymen te vormen en bij sommige zaden met kiemrust (dormantie) kan het toevoegen van gibberellinezuur de kiemrust breken. Het is een van de eenvoudigste gibberellines.
De naam gibberelline is afgeleid van de schimmelnaam Gibberella fujikuroi. Deze schimmel werd in 1926 door de Japanner Eiichi Kurosawa ontdekt als de veroorzaker van de bakanae-ziekte (domme zaailing ziekte) in rijst. De door de schimmel gevormde gibberelline veroorzaakte een sterke plantengroei waardoor slappe planten met lange internodiën (leden) ontstonden. Enkele jaren later werd door Japanse onderzoekers uit de zieke planten twee kristallijne, biologisch werkzame stoffen geïsoleerd. Deze werden gibberelline A en gibberelline B genoemd. Pas vele jaren later is de werking van gibberellinen pas goed begrepen door het toentertijd gestarte onderzoek van het United States Department of Agriculture (USDA) en door de Imperial Chemical Industries (ICI) in de Verenigde Staten. Tijdens dit onderzoek werden de nieuwe gibberelline X en gibberellinezuur geïsoleerd. Later bleek gibberelline X een mengsel te zijn van gibberelline A en gibberellinezuur. Bij verdergaand onderzoek in Japan bleek gibberelline A te bestaan uit de drie verwante stoffen gibberelline A1, gibberelline A2 en gibberelline A3. Gibberellinezuur bleek gelijk te zijn aan GA3.
Er zijn meer dan 110 verschillende gibberellines (GA1 ... GA110), waarvan er slechts enkele in planten actief zijn, zoals GA1, GA3 en GA7.
Zeer lage concentraties hebben al een groot effect, maar te hoge concentraties werken juist remmend.
Hoe wordt het toegepast?
Stap 1)
Plant zoveel mogelijk plantjes. Je kan het met 20 plantjes proberen, maar als je een effectief resultaat wilt boeken, zal je het met een 1000tal moeten doen.
Stap 2)
Laat je plantjes wat groeien. Pakweg een weekje of twee laten groeien. Zeker nog niet in bloei zetten! Geen 12/12 cyclus maar een gewone groei cyclus zoals 18/6!
Stap 3)
Onderzoek je plantjes een voor een onder de microscoop. De sleutel hier is geduld. Bij ieder plantje zoveel mogelijk cellen bekijken en kijken of er XY chromosomen bij zitten.
Zo ziet een XY chromosomenpaar er uit:
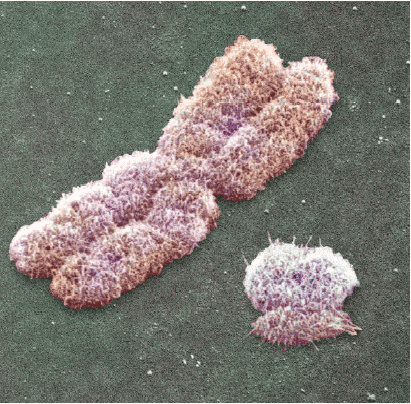
Deze planten moet je apart zetten. Het doel is dat je op zoek gaat naar een plant met 100% XX chromosomenparen (dit zijn de vrouwelijke)
XX => vrouw
XY => man
Als je deze plant gevonden hebt (uit de 1000 plantjes ga je er nog geen vinden die 100% vrouwelijk is, maar rond de 99% zou wel moeten lukken. 100% is theoretisch mogelijk maar in de praktijk niet) dan ga je nr stap 4.
Stap 4)
Zet je 100% vrouwelijke plant in bloei. Als het bloeimoment juist is (dwz dat het moment van optimale bevruchting bijna is aangebroken) dan spuit je een oplossing Gibberrelinezuur van 0,00X per liter water.
De 0,00X oplossing is dus een oplossing van 0,00x g Gibberrelinezuur in een liter water. De X is onbekend. Hier moet je zelf mee experimenteren.
Als ik afga op wat ik al gelezen heb hierover kan je best de gulden middenweg nemen en gewoon 0,005 g per liter doen.
Als iemand informatie heeft over nauwkeurigere verhoudingen zou dit wel interessant zijn.
De theorie achter dit zuur:
Gibberrelinezuur zorgt ervoor dat er een verandering in hormoonsverhoudingen optreed.
Kort gezegd:
De buds van je plant worden beïnvloed door dit zuur en veranderen hun verhouding van 99% vrouwelijk naar ... % mannelijk.
Deze mannelijke bloemen zorgen ervoor dat de plant zelf bevrucht wordt.
Doordat de plant 99% vrouwelijk is, zal hij ook 99% vrouwelijke zaadjes produceren.
zie: Produceren van vrouwelijke zaden
Vrouwelijke zaden kweken
Gefeminiseerde / vrouwelijke zaden creëren is een kunst, er zijn enkele verschillende manieren. Je hebt glibberic Acid, licht stress, ph-stress, stress door je meststoffen maar allemaal willen ze een ding bereiken dat er mannelijke bloemen op een XX plant komen.
Dit proces kan toegepast worden om:
A: nieuwe vrouwelijke zaden te creëren van een supermoeder die je staan hebt
B: om interessante vrouwelijke hybride zaden te creëren van verschillende interessante soortjes die je momenteel hebt
C: om vrouwelijke zaden te creëren voor een optimale buitenkweek
D: om het proces te versnellen naar de zoektocht voor een ideale nieuwe moederplant
E: om het aantal planten die je kweekt voor medicinaal gebruik zo beperkt mogelijk te houden (dus geen extra planten uit angst voor teveel mannetjes)
F: voor meer variatie, door stabiele vrouwelijke zaadlingen te maken die kunnen dienen als een alternatief voor klonen
Het is wel een vereiste dat je je al goed ingestudeerd hebt over de theorie van cannabis kweken voor dat je een techniek als deze gebruikt.
Het is ook zeer belangrijk dat je de basis veiligheidsvoorschriften respecteert wanneer je de chemicaliën mixt en gebruikt. De risico`s zijn gelijk aan het mixen en gebruik van chemische meststoffen dus pas dezelfde veiligheids voorzorgen toe.
Onthoud: niets zal nooit goeie genetica vervangen, en een deel van wat je kweekt zou altijd moeten terugkeren op de professionele cannabis kweker... Diegene die vele generaties gewerkt heeft om van een soortje uiteindelijk hun echte F1 meesterwerkjes te bekomen. Help hen door hun zaden te kopen zo kunnen zij verder werken aan nieuwe sterke soorten voor ons thuiskwekers..
-------------------------------------
1 STS
Preparation of STS:
Eerst word een voorraad oplossing gemaakt. Het bestaat uit 2 delen (A en B) die eerst apart worden vermengd en uiteindelijk dan samen vermengd.
Deel A wordt altijd samen gemixed met deel B terwijl je snel roert. Gebruik gedestilleerd water, kraantjeswater kan precipitaten vormen.
Draag handschoen tijden het mixen en gebruiken van deze chemicaliën, en mix en gebruik deze enkel in een deftig geventileerde ruimte. Een masker kan voorkomen dat je een bepaalde stof inademt, die agressief is. STS is kleurloos en geurloos, en betekent een minimum aan risico als je alles toepast zoals hier boven beschreven. Onthoud dat silvernitraat en sts bruine plekken kunnen veroorzaken bij het indrogen, dus spuit over krantenpapier en vermijd morsen.
Deel A: 0.5 gram zilver nitraat vermengd met 500ml gedestilleerd water
Deel B: 2.5 gram natriumthiosulfaat (watervrij) vermengd met 500ml gedestilleerd water
Het zilvernitraat lost op binnen de 15 seconden. Het natriumthiosulfaat heeft ongeveer 30/45 seconden nodig om op te lossen.
De zilvernitraat oplossing (deel A) word dan vermengd met het natriumthiosulfaat oplossing (deel B) terwijl je goed en snel roert. Het resultaat is een voorraad zilver thiosulfaat oplossing (STS).
Deze voorraad oplossing wordt vervolgens verdund in een verhouding van 1:9 om een werkende oplossing te maken. Bijvoorbeeld: 100ml STS is toegevoegd aan 900ml gedestilleerd water. Deze oplossing wordt dan verneveld op de geselecteerde vrouwelijke planten.
De beide producten de voorraad STS zowel als de gemaakte oplossing word best in de koelkast bewaart na gebruik, om activiteits verlies te voorkomen. Teveel aan oplossing kan altijd veilig door het wc gespoeld worden of enige andere afvoer met stromend water met verwaarloosbare gevolgen voor het milieu.
-------------------------------------
Gebruikswijze:
De STS oplossing wordt dan besproeid op de vrouwelijke planten tot het er mooi afdruipt. Doe het sproeien over een krant in een aparte ruimte. Je zal waarschijnlijk niets ruiken maar ventileer toch maar. Nu heb je wat ik een "F>M plant" noem; een vrouwelijke plant die mannelijke bloemen gaat produceren.
Nadat de F>M plant droog is, plaats je ze direct onder 12/12. Dit wordt meestal 3/4 weken voor de doel datum (om te bestuiven) wanneer de planten klaar zijn om bestuift te worden. Responstijden kunnen enigszins variëren, afhankelijk van de soort. Meer specifieke tijden kunnen worden bepaald door met uw eigen strain proeven te doen. In mijn onderzoek duurde het 26 dagen voor de eerste pollen. 30-35 dagen lijkt een optimale planning.
Zo, veronderstel dat je doelplant 3/4 weken nodig heeft om volledig volwassen zaden te produceren, een strain dat 8 weken nodig heeft om volwassen te worden moet in bloei gezet worden op ongeveer dezelfde tijd als de F>M plant. Een doelplant dat afbloeit in 6weken moet dan 10 dagen later in bloei gezet worden zodat ze niet al afgebloeid is voor de zaden volledig volwassen zijn.
Een afzonderlijke tak die je bestoven hebt mag je altijd iets langer volwassen laten worden aan een plant, je moet ze niet meeoogsten samen met de zaadloze toppen. Laat enkel genoeg bladeren aan de plant zodat ze gezond kan blijven.
-------------------------------------
Effecten:
In enkele dagen zag ik al bladeren vergelen aan de F>M planten. Dit verschijnsel bleef ongeveer 2 weken duren: daarna werden ze langzaam terug groen, buiten enkele grote fan bladeren. Voor de rest leek de plant gezond. Geen verbranding te zien. De groei stopte wel voor 10 dagen, en starte erna terug langzaam. Geen strekken te zien. Na 2 weken vormden de F>M planten duidelijke mannelijke bloemen. Niet enkel een paar ballen maar echte complete mannelijke toppen. 1 plant vormde nog vrouwelijke bloemen, maar in het geheel bekeken was deze overwegend mannelijk.
Het is vreemd om een oude vriendin die je als de achterkant van je hand kent te zien een seksverandering te ondergaan.
Als de F>M planten eruit zien alsof zij snel kunnen openen pollen laten vrijkomen, (3-1/2 tot 4 weken) verplaats je hen best naar een aparte ruimte (weg van je bloeiruimte) met licht op 12/12. Je moet niet teveel inzitten over wattages; het is maar tijdelijk.
Als de pollen vrijkomen verplaats je best je doelplanten bij in de kast zodat ze deze kunnen bestuiven.
Een meer gecontroleerde aanpak is je F>M planten te isoleren in een derde afzonderlijke ruimte(geen licht is hier nodig, deze zijn nu toch pollen aan het lossen dus zijn ze toch op hun einde). In deze andere ruimte kunnen we de pollen voorzichtig verzamelen met een plastiekzak of een vel krantenpapier. Dan kunnen we onze doelplanten in de bloeiruimte op deze manier bevruchten. Als je het zo doet let wel op dat je pollen van je F>M plant niet mixen of dat ze niet kruis bestuiven. Dus vermijd beweging.
Let er speciaal op dat deze pollen zich niet verzamelen buiten aan de buitenkant van je zak door statische elektriciteit bv.
Neem kleine open clusters van de binnen kant van de bloemen in de zak en schud ermee. Ga dan best naar buiten om de overtollige lucht uit de zak vrij te laten. Let hierbij op dat er geen pollen op je kleren of handen terecht komen.
Nu kan je deze zak voorzichtig over een tak schuiven en dan afbinden rond de tak met een stukje tape of koord, zodat de pollen bij de top opgesloten zit. Laat de zak zich langzaam opvullen met een beetje lucht voor je hem afsluit aan de tak, zodat de tak nog kan "ademen". Deze techniek zorgt ervoor dat je bij een plant verschillende takken kan bestuiven met verschillende pollen van verschillende strains of dat je een deel van de plant ook nog kan oogsten als sensemilla.
Na 4-5 dagen mag je de zak voorzichtig verwijderen en de plant kan zen bloeicyclus verder vervolledigen.
Maar gelijk welk van deze wijzes dat je toepast uiteindelijk heb je na 2/4weken rijpen vrouwelijke zaden over.
-------------------------------------
Over de chemicalieen:
Zilvernitraat is een witte kristallijne lichtgevoelige chemische stof die gewoonlijk wordt gebruikt in de fotografie. Het wordt ook gebruikt in de ogen van baby`s bij de geboorte om blindheid te voorkomen. Het kan leiden tot lichte irritatie van de huid, en bruine vlekken. Vermijd inademen. Ik heb geen geur of rook ondervonden maar ventilatie word aanbevolen. Een halve gram STS kost verrassend weinig en zou passen in een gel-capsule.
Have fun met het testen van deze techniek. Gebruik het met verstand. En bovenal gebruik STS bij kwaliteits F1 zaden geproduceerd door een professionele kweker.
2 Rodelization (soma)
In mijn zoektocht naar mooiere milieuvriendelijke manieren voor te werken met cannabisplanten, heb ik een nieuwe manier ontdekt om vrouwelijke cannabiszaden te ontwikkelen. Vrouwelijke zaden ontstaan door stressen niet door genetica. Alle cannabisplanten kunnen en zullen mannelijke bloemen produceren bij stress. Sommige soorten hebben liever een hogere P, sommige een lagere. Sommigen hebben veel voeding nodig anderen dan weer minder. Daar zit veel variëteit in de cannabis genetica, en je kan niet elke plant gelijk behandelen.
Als je een bepaalde soort wilt leren kennen neemt het echt een groot aantal kweken in beslag voor je je echt kan zeggen dat je die soort kent. net als je een vriend wil leren kennen, dit neemt veel tijd in.
Ik groei nu al verschillende strains voor meer dan een decennium en ik begin echt alle nuances te kennen die verschillende planten bezitten. Ik kan ze herkennen van op een afstand. Ik moet wel zeggen dat ik hierbij veel hulp krijg van mijn vrienden, zowel voor het maken van zaden als voor het beter leren kennen van deze heilige plant.
Ik noemde deze nieuwe methode 'Rodelization' maar een vriend die me hielp te realiseren en gebruik te maken van deze manier om vrouwelijke zaden te maken. Nadat ik dezelfde soort kweekte oogst na oogst onder dezelfde condities, merkte ik dat als ik deze planten 10 tot 14 dagen langer liet staan dan gewoonlijk dat deze mannelijke "bananen" vormde. Een mannelijke banaan is een zeer kleine mannelijke bloem die zich vormt door stress. Normaal laten zij geen pollen vroeg genoeg los om cannabis zaden te maken, maar soms wel. Dit is een soort ingebouwde veiligheid voor de plant, zodat de plant zeker is dat zij zich kan voortplanten ook in extreme condities.
Voor mij is een mannelijk bloem een zeer mooi gebeuren. Het geeft de mogelijkheid om vrouwelijk zaad te maken. Veel kwekers hebben een natuurlijke angst voor mannelijke bloemen 'bananen'. Ze zien een bloem en willen al direct hun hele oogst weggooien of gaan er met een pincet angstwekkend rond de planten om deze banaantjes te plukken. Ik noem deze 'veiligheids voorzieningen' omdat deze voorkomen bij stress.
Gebruikswijze:
Bij de Rodelizaion methode is de mannelijke banaan zeer waardevol. Nadat je je vrouwelijke plant 10/14 dagen langer dan normaal hebt laten bloeien, hang ze op om te drogen, neem ze nadien voorzichtig van de drooglijnen en inspecteer voor bananen. Elke banaan zou moeten verwijdert worden en voorzichtig in een kleine zak geplaatst worden. Deze afgesloten zakken kan je in de frigo bewaren voor 1 tot 2 maand en ze zullen nog steeds potent zijn.
Voor de tweede fase heb je een oogst nodig dat al 2.5 weken in bloei staat. Neem je gesloten zak uit de frigo, en vervolg je plan om de vrouwelijke toppen te bevruchten. Om dit te doen, moet je eerst de vrouwelijke plant en de pollen van de vorige strain uitkiezen. Dan zet alle vents en fans uit in de ruimte. Neem een kleine verfborstel, dip het in de pollen, en verf het op de vrouwelijke toppen. Doe dit voor iedere strain die je wil samen groeien. Ik heb dit gedaan met 10 verschillende strains in dezelfde ruimte met groot succes.
Ik gebruik voornamelijk de lager gelegen takken om weed zaad van te maken, de topcola`s laat ik zaadloos om te roken. Deze methode neemt wel tijd in (2oogsten), maar is volledig organisch en zorgt ervoor dat je mooie kwaliteit rookbare weed ook hebt en tegelijkertijd maak je ook vrouwelijke zaden aan. Als je een van die kwekers bent die nooit zaden heeft durven kweken uit angst om niets goed over te hebben om te roken, dan zal je zekers houden van deze manier van werken.
Je kan natuurlijk ook deze pollen gebruiken om nieuwe kruisingen vrouwelijke zaden te maken, door kruisbestuiving. De oudere vrouwen met de banaantjes kan je ook in de kamer met de jongere vrouwtjes zetten als ze ongeveer 3 weken in bloei zijn. Zet de circulatie fans vollen bak, en de kleine beetjes pollen vinden hun weg wel terug door de kweekruimte. Houdt dit enkele dagen vol en 6 tot 7 weken later heb je 100% rijpe vrouwelijke zaden; niet zoveel als er zouden gemaakt zijn moest er een mannelijk plant bijgestaan hebben maar genoeg om opnieuw te starten ergens met dezelfde genetica.
Soma
3 Ethyleen gas
Behandelen van cannabis zaad met ethyleen gas zorgt ervoor dat de kans op meer vrouwelijke planten verhoogt met ongeveer 50% (volgens sommigen zelfs 100%, wil ik wel es testen desnoods met hempzaad). Ethyleen word geproduceerd door verschillende planten (bv. bananen, komkommers en meloenen), en deze kunnen op een gemakkelijke manier gebruikt worden om cannabiszaad te behandelen.
Gebruikswijze:
Ongeveer 2 weken voor je van plan bent om je zaadjes te ontkiemen, steek ze in een papierzak of envelop en steek deze in een plastiek zak samen met de bananenschillen van een nog rijpende banaan of komkommer. Vervang de schillen om de paar dagen samen met de papieren zak zodat je schimmel voorkomt.
4 Colloïdaal Zilver
Wat is Colloïdaal Zilver?
Colloïdaal zilver is zuiver, metallic zilver (het element), met deeltjes van 15 atomen of minder, elk met een positieve elektrische lading en bevestigd aan een molecuul van een eenvoudig eiwit. Deze elektrisch geladen deeltjes van zilver zijn uiterst klein, meestal variërend van ongeveer 0.001 tot ongeveer 0,01 micron in diameter, en zijn gesuspendeerd in gedeïoniseerd water. De kracht van de elektrische lading is sterker dan de kracht van de zwaartekracht, dus de zilveren deeltjes blijven opgeschort.
Hoe wordt het toegepast?
De zilverionen in colloidaal zilver reageert als ethyleen remmer/antagonist die de plant forceert om mannelijke bloemen aan te maken die pollen bevatten die enkele vrouwelijke chromosonen bevatten.
Quote:
Treatments which reduce the ethylene level in the tissues (hypo-baric conditions, treatment with benzothiodiazole) or antagonize the action of ethylene (CO2) cause the formation of male or bisexual flowers in place of female ones (Byers et al. 1972)
Wij gebruiken eenvoudigweg een normale lege spuitbus om het op de bladeren te sproeien van een geweten vrouwelijke plant om de pollen op te wekken. Dit kan je doen Met als resultaat 'gefiminiseerde pollen' in tegenstelling tot het regulier stuifmeel bij een man. Als gevolg hiervan, is elk vrouwtje bestoven met de gefeminiseerd pollen een producent van uitsluitend vrouwelijk zaad. We kunnen hier zelfs een moeder voor gebruiken van een 'CLONE-ONLY' strain, zelfs een vrouwelijke plant gegroeid uit feminised zaad. Het enige dat van belang is dat dit een vrouwelijke plant is.
aanrader om te lezen
Induction of Fertile Male Flowers in Female Cannabis (Uni paper on feminised seeds)
H.Y. Mohan Ram and R. Sett. Department of Botany, University of Delhi, Delhi (India)
Beetje uitgebreider:
-Wanneer start ik best met sproeien
Je begint best 2 weken voor de planten op 12/12 gaan en blijven sproeien tot je pollen ziet ontwikkelen.
-Hoe lang sproeien
Normaal gezien sproeien we ongeveer 2 weken.
(eenmaal de plant besproeit, is ze niet langer rookbaar. Dus niet oproken!!)
-Je kan aan zelfbestuiving doen
Dit gebeurt ook in de natuur bv. als een plant zeer oud word (rhodelization), waarbij een vrouwelijke plant enkele bananen afgooit om zichzelf te bestuiven. Dit is een soort van species survival mechanism veronderstel ik.
Je verkrijgt wel niet veel zaad op deze manier en sex tussen twee personen is de norm in het leven, dus persoonlijk probeer ik altijd een 2e vrouwtje als doel voor de mannelijke pollen in de ruimte te plaatsen, en gebruik ik zelfbestoven planten enkel als ik geen andere optie heb.
Ik vermoed dat mss de zelfbestoven zaden kunnen uitgroeien als een kloon-a-like? Maar daar ben ik niet zeker van.
-Op welke mogelijke manieren kan dit allemaal falen
Collodiaal Zilver is zo makkelijk om te maken, gebruiken, en het geeft ons de mogelijkheid om zelfs de nmoeilijkste soortjes te forceren, dus het volledige proces is eigenlijk zeer eenvoudig en zeer effectief als je de basis volgt.. Maar als je dit niet doet, kan het volledig verkeerd zijn.Als je ooit al es CS gebruikt heb maar het niet gelukt is om een plant te reversen, dan is er een goeie kans dat je simpelweg een foutje gemaakt hebt in de procedure.
-De hoofdproblemen
- geen gedestilleerd water gebruiken
- geen zuiver zilver gebruikt 999.999
- de elektroliese niet lang genoeg laten duren
-sproeien met een niet sterk genoege oplossing
-niet genoeg gesproeid (de plant moet volledig doorweekt zijn)
-niet regelmatig genoeg gesproeid
-niet lang genoeg gesproeid (sommige soortjes moet je wel es 3 weken sproeien)
Hoe maak je colloidaal zilver?
Benodigdheden voor de colloidaal zilver generator:
1)een DC adapter (9V/ 300mA-800mA).(600mA is optimaal) gedestilleerd water
2)1/2 quart mayonnaise bokaal 2x 99.99% pure zilver draden , munten, of een zilver bar
3)aquarium luchtpompje
4)2 geisoleerde alligator clips
5)houten plankje van 10cm x 20cm
6)zaag of zwitsers zakmes
7)houtlijm
8)elektrische boormachine en bit
9)een plastiek deksel voor de mayonaise pot of een rubber elastiek ofzo
10)een stukje hout
Maken van de generator
1)Zaag je houten plankje zodanig dat de bokaal en de adapter erop passen
2)Zaag het ander houten latje zodanig dat je het van boven over de mayo pot kan plaatsen. Boor 2 gaten 2 cm van elkaar (om de zilverdraden op te houden). Die gaten zouden iets groter moeten zijn dan de diameter van de zilverdraden. Omdat we daar dan die draden gaan doorsteken. Lijm 2 smalle houten plankjes op de uiteinden van het houten latje, zodat het latje past op de bovenkant van de mayonaise pot
Indien je zilver stukken of bars gebruikt, moet je dit design wel een klein beetje aanpassen.
3)Knip bij de AC adapter de connector af, niet te kort zodat je toch nog beetje draad over hebt. Strip een 2 tal cm draad af
4)Bevestig de gestripte draad aan de aligatorklemmetjes
5)Monteer de generator zoals op de foto
6)Vul de bokaal met gedestilleerd water tot een paar cm van de top
7)Sluit de 2 alligator klemmen aan op de zilver draden die uitsteken uit de houten lat, en steek ze door de geboorde gaten in de houten lat op de bokaal.
8)Sluit de adapter en luchtpomp aan op een timer. Nu ben je klaar om de CS oplossing te maken.
9)Zet de timer op 8 tot 20uren (8u zou genoeg moeten zijn, maak dat je +30PPM hebt in je oplossing
10)Let erop dat de pomp en de ac adapter werkt. De pomp kan je horen maar de DC output ga je moeten testen met een voltmeter. De bubbels zorgen ervoor dat de kwaliteit verbeterd van de zilver oplossing. Kleine fuzzy bubbels zullen zich vormen in 1 pool. Een zaklamp kan je helpen om dit te zien.
11)Nadat dit proces afgelopen is, zeef je je oplossing door een niet gebleekt koffiefilter om kristallen en witte schilfers te verwijderen. Bewaar het elexir in een donkere glazen fles op een koele plaats. Je kan deze enkele maanden bewaren maar het is wel best dat je kleine hoeveelheden maakt zodat je het steeds vers kan gebruiken.
12)Was de mayonaise bokaal en luchtpomp na ieder gebruik en dan spoelen met gedestilleerd water. Reinig de zilverdraden met een scotchbrite pad tegen onzuiverheden en zilverkristallen, spoelen erna.
13)Bewaar nadien alles samen op een stofvrije plaats.
nog enkele manieren om een generator te maken + gebruikswijze: http://www.quantumbalancing.com/makeyourowncs.htm
Waarom CS gebruiken?
Goed, u kunt ook gebruik maken van zilver theosulphate (= nitraat + natrium theosulphate), zilvernitraat (zilver + salpeterzuur), of gibberellinezuur, die allemaal succesvol zijn gebleken in deze taak, maar CS is niet giftig , niet-bijtende, gemakkelijk en veilig en goedkoop te maken thuis, vereist geen DEA aanvraagformulier, en is niet een gereguleerde stof. U kunt het zelfs kopen bij uw plaatselijke drogist / apotheek (sommige mensen drinken het voor haar veronderstelde voordelen voor de gezondheid), en het is goedkoop.
Voor mij, is colloïdaal zilver ongeëvenaard. Colloïdaal zilver werkt als ethyleen remmer / antagonist, wat ons toelaat om het proces te forceren op elk moment we kiezen, zelfs op hard stress-resistente planten (CS maakt geen gebruik van stress om zen magie uit te voeren).
5 Gibberellinezuur
Wat is Gibberellinezuur?
Gibberellinezuur (GA3) is een plantenhormoon, dat de groei en strekking van plantencellen bevordert. Ook beïnvloedt het de verhouding tussen mannelijke en vrouwelijke bloemen op een plant, bevordert het de vruchtzetting, geeft het grotere bladeren en kan sommige tweejarige planten al in het eerste jaar laten bloeien. Verder stimuleert het de cellen van kiemende zaden om mRNA coderend voor hydrolytische enzymen te vormen en bij sommige zaden met kiemrust (dormantie) kan het toevoegen van gibberellinezuur de kiemrust breken. Het is een van de eenvoudigste gibberellines.
De naam gibberelline is afgeleid van de schimmelnaam Gibberella fujikuroi. Deze schimmel werd in 1926 door de Japanner Eiichi Kurosawa ontdekt als de veroorzaker van de bakanae-ziekte (domme zaailing ziekte) in rijst. De door de schimmel gevormde gibberelline veroorzaakte een sterke plantengroei waardoor slappe planten met lange internodiën (leden) ontstonden. Enkele jaren later werd door Japanse onderzoekers uit de zieke planten twee kristallijne, biologisch werkzame stoffen geïsoleerd. Deze werden gibberelline A en gibberelline B genoemd. Pas vele jaren later is de werking van gibberellinen pas goed begrepen door het toentertijd gestarte onderzoek van het United States Department of Agriculture (USDA) en door de Imperial Chemical Industries (ICI) in de Verenigde Staten. Tijdens dit onderzoek werden de nieuwe gibberelline X en gibberellinezuur geïsoleerd. Later bleek gibberelline X een mengsel te zijn van gibberelline A en gibberellinezuur. Bij verdergaand onderzoek in Japan bleek gibberelline A te bestaan uit de drie verwante stoffen gibberelline A1, gibberelline A2 en gibberelline A3. Gibberellinezuur bleek gelijk te zijn aan GA3.
Er zijn meer dan 110 verschillende gibberellines (GA1 ... GA110), waarvan er slechts enkele in planten actief zijn, zoals GA1, GA3 en GA7.
Zeer lage concentraties hebben al een groot effect, maar te hoge concentraties werken juist remmend.
Hoe wordt het toegepast?
Stap 1)
Plant zoveel mogelijk plantjes. Je kan het met 20 plantjes proberen, maar als je een effectief resultaat wilt boeken, zal je het met een 1000tal moeten doen.
Stap 2)
Laat je plantjes wat groeien. Pakweg een weekje of twee laten groeien. Zeker nog niet in bloei zetten! Geen 12/12 cyclus maar een gewone groei cyclus zoals 18/6!
Stap 3)
Onderzoek je plantjes een voor een onder de microscoop. De sleutel hier is geduld. Bij ieder plantje zoveel mogelijk cellen bekijken en kijken of er XY chromosomen bij zitten.
Zo ziet een XY chromosomenpaar er uit:
Deze planten moet je apart zetten. Het doel is dat je op zoek gaat naar een plant met 100% XX chromosomenparen (dit zijn de vrouwelijke)
XX => vrouw
XY => man
Als je deze plant gevonden hebt (uit de 1000 plantjes ga je er nog geen vinden die 100% vrouwelijk is, maar rond de 99% zou wel moeten lukken. 100% is theoretisch mogelijk maar in de praktijk niet) dan ga je nr stap 4.
Stap 4)
Zet je 100% vrouwelijke plant in bloei. Als het bloeimoment juist is (dwz dat het moment van optimale bevruchting bijna is aangebroken) dan spuit je een oplossing Gibberrelinezuur van 0,00X per liter water.
De 0,00X oplossing is dus een oplossing van 0,00x g Gibberrelinezuur in een liter water. De X is onbekend. Hier moet je zelf mee experimenteren.
Als ik afga op wat ik al gelezen heb hierover kan je best de gulden middenweg nemen en gewoon 0,005 g per liter doen.
Als iemand informatie heeft over nauwkeurigere verhoudingen zou dit wel interessant zijn.
De theorie achter dit zuur:
Gibberrelinezuur zorgt ervoor dat er een verandering in hormoonsverhoudingen optreed.
Kort gezegd:
De buds van je plant worden beïnvloed door dit zuur en veranderen hun verhouding van 99% vrouwelijk naar ... % mannelijk.
Deze mannelijke bloemen zorgen ervoor dat de plant zelf bevrucht wordt.
Doordat de plant 99% vrouwelijk is, zal hij ook 99% vrouwelijke zaadjes produceren.
zie: Produceren van vrouwelijke zaden



 (a) The possible genetic changes that could occur in the transition from cosexual to separate sexed populations. (b) Effects of a female-sterility 'modifier' allele on hermaphrodites and females, in the absence of sex-limitation of its phenotypic effects. A trade-off between male and female functions is assumed, so that a gene increasing male fertility will often have the effect of reducing female fertility.
(a) The possible genetic changes that could occur in the transition from cosexual to separate sexed populations. (b) Effects of a female-sterility 'modifier' allele on hermaphrodites and females, in the absence of sex-limitation of its phenotypic effects. A trade-off between male and female functions is assumed, so that a gene increasing male fertility will often have the effect of reducing female fertility.





Comment